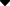


为了更好地理解神经毒素对神经肌肉传递的阻断作用,有必要对神经肌肉之间正常的信号传递过程做一个概括性的介绍。
(―) 细胞的静息电位
静息电位是指细胞在不受刺激时,细胞内外的电位差。由于这种电位差存在于静息细胞膜的两侧,故又称跨膜静息电位,简称静息电位或膜电位。只有可兴奋细胞(如肌肉细胞和神经细胞)才具有静息电位。大多数细胞的静息电位是一个稳定的直流电位,但各种细胞的静息电位值是不同的,如乌贼的巨型轴突和青蛙的骨骼肌细胞的静息电位为-50mV^-70mV,即膜内电位比膜外电位低50mV70mV,哺乳动物的肌肉和神经细胞为-70mV90mV。
静息电位的产生与细胞膜内外离子的分布和运动有关。正常情况下,细胞内K+浓度和有机阴离子A-浓度高于膜外,Na+和Cr浓度高于膜外高于膜内。在这种情况下,K+和A-必然有向膜外扩散的趋势,但细胞膜在安静时对K+的渗透性更强,而对有机负离子A-没有渗透性。因此,K+通过开放的K+通道从膜内沿浓度梯度扩散到膜外。随着K+ 扩散到膜外,膜外带有更多的正电荷。同时,有机负离子不能通过膜。留在膜中。这导致膜外侧变为正极,膜内侧变为负极,膜内外出现电位差。 K+扩散到膜外引起的电位差,内负外正,成为阻止K+向外移动的力,最终达到平衡,形成稳定的膜内外电位差。这种内负外正的状态称为极化状态。
(2)动作电位及其传导
当可兴奋细胞受到刺激时,它们都有一个共同的、较早的反应,即在受刺激的细胞膜两侧发生快速、可逆的电变化,称为动作电位。当神经或肌肉细胞在安静的情况下受到短阈值或超阈值刺激时,膜内原来70mV-90mV的负电位会迅速消失,然后变成+20mV~+40mV的正电位,即,由原来静止时,内负外正变为内正外负,电位变化幅度为90mV~130mV。这个过程称为去极化。其中,膜内电位由零变为正的过程称为反向极化。上述刺激引起的变化只是暂时的,膜两侧电位迅速恢复到静止状态和水平。这个过程称为复极化。神经纤维的动作电位一般只持续0.5ms~20ms。如果描成曲线,会呈现尖锐的脉冲,称为尖峰,包括快速上升支(即去极化相)和快速下降支。 (即复极相)。在尖峰完全恢复到静息电位水平之前,膜电位会发生缓慢的波动,称为后电位。
用直流电刺激神经纤维的实验发现,通电时负极下的膜发生去极化,出现动作电位。这是因为通电时,负极下方的隔膜有向外的电流通过。由于膜具有一定的电阻值,因此在膜的两侧产生内部正和外部负压降。电位方向相反,使静息电位值降低,即膜处于一定程度的去极化状态。当膜电位降低到某一临界值(这个临界值比静息电位小10mV~20mV左右,称为阈值电位)时,膜上大量Na+通道被激活,膜的通透性下降。膜对Na+ 的突然增加,并暂时超过膜对K+ 的通透性。在膜两侧Na+浓度差和静息电位的驱动下,大量Na+快速内流,膜进一步去极化,导致更多Na+通道开放。在反馈作用下,更多的Na+快速流入。随着正电荷的进入,膜中的负电势迅速消失,然后变为正电。当膜中的正电位增加到足以阻止浓度差驱动的Na+ 内流时,Na+ 内流变小。膜中的电位不会停留在正电位状态,而是会很快发生复极化。钠通道开放时间很短,很快进入所谓的失活状态,因此膜对Na+的通透性变小。同时,膜对K+的通透性增加并逐渐超过对Na+的通透性,因此膜内的K+在浓度差和电位差的驱动下向膜外扩散,使膜内电位从正值向负值变化,直到恢复静息电位水平。
当某一节段产生动作电位时,相邻的神经节段仍处于内负外正的极化状态。因此,在激发段和相邻的未激发段之间存在电位差。由于电位差的存在,电荷发生运动,形成局部电流,电流使未激发部分的膜去极化。当去极化达到阈电位水平时,立即产生动作电位,即相邻的未兴奋节段开始兴奋。状态,继续如此,实现兴奋对神经的传导。
(3) 神经肌肉连接处的兴奋性传递
支配骨骼肌的运动神经源自中枢神经系统内的运动神经元。当运动神经元的轴突接近肌肉时,它会失去髓鞘并分叉。一般来说,每个分支支配一根肌纤维,因此,一个运动神经元可以支配多根肌纤维。每根神经纤维末端与受神经支配的肌纤维之间没有结构连接,两者之间有20nm的间隙。这个间隙与一般的细胞外液相通。这种结构称为神经肌肉接头,与神经元之间的化学突触非常相似。轴突末端嵌入肌细胞膜的凹陷处,轴突末端的膜称为前关节膜。肌肉细胞膜的凹陷部分称为关节后膜,或终板,终板膜向肌浆凹陷成许多小褶皱,这可能意味着关节后膜的面积增加。轴突末端含有大量的突触小泡(如图5-20-2所示)。突触小泡是直径约50nm、无特殊结构的囊泡,内含乙酰胆碱,乙酰胆碱在轴浆中合成并贮存在囊泡中。
当神经纤维处于安静状态时,一般只有少量小泡随机靠近轴突末端,小泡中的乙酰胆碱通过胞吐作用释放到关节间隙中。由于仅释放少量乙酰胆碱,因此对肌肉细胞没有显着影响。但当神经纤维向神经末梢传递兴奋时,在动作电位去极化相的影响下,轴突末梢膜上的Ca2+通道打开,细胞外液中的一部分Ca2+进入膜内,引起大量突触小泡被释放。乙酰胆碱与终板膜上的乙酰胆碱受体结合,这种结合导致受体蛋白分子
构象改变,这种改变使 终板膜对离子(主要是Na+)的通 透性增加,结果使终板膜去极化,这 一电变化称为终板电位。终板电位 是局部兴奋,当肌细胞膜的去极化 达到阈电位水平时,就产生一次动 作电位,此动作电位向整个肌细胞 进行“全或无”式的传导,这就是神 经-肌肉接头的一次兴奋传递过程。一次神经冲动引起末梢释方文乙醜月旦目5-20-2神经-肌肉接头的结构碱量足以引起肌细胞产生一次动作电位。乙酰胆碱在完成上述作用后,被胆碱酯酶迅速破坏。因此,一次神经冲动只能引起 肌细胞一次兴奋,诱发一次收缩。
前面已经说过,在没有神经冲动的静息状态下,仍然有小量的突触囊泡随机释放,这 样其内的乙酰胆碱可以与终板的受体结合形成微终板电位(mEPP)。微终板电位与正常的终板电位(EPP)有相同的形状,但振幅仅0. 5mV,它们不足以引起肌肉收缩。微终板电 位只有在终板区才能找到。它们的出现是随机的,但一般也有一个大致的频率。微终板电 位的值也是大小不等的,但也有一个大致的平均值。通过测定电位的大小可以计算出突触 小泡释放乙酰胆碱的量,也可以计算出电荷的流量,这种计算对终板电位和微终板电位都 是适用的。
(四)骨骼肌的收缩
终板电位引起横管膜电变化,这种变化使邻近的终池膜结构中某些带电基团的移位, 从而使位于终池膜的Ca2+通道开放,于是Ca2+就顺着浓度差由终池向肌浆中扩散,导致 肌浆中的Ca2+浓度升高。进入肌浆中的Ca2+弥散到肌原纤维周围,在那里与肌钙蛋白结 合,引起肌肉收缩。只要肌浆中的Ca2+处于高浓度,肌肉收缩就将继续。然而,存在于肌质 网膜结构中的特殊蛋白质-Ca2+泵不断地将肌浆中的Ca2+在逆浓度差的情况下转运回终 池内贮存,于是肌浆中的Ca2+浓度很快又降低,肌肉又松弛。
















