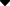


蛇毒神经毒素是蛇毒中毒性较大的成分,可使动物出现弛缓性麻痹和呼吸衰竭,导致动物死亡。这种毒素广泛存在于眼镜蛇和海蛇的毒液中,只有某些种类的响尾蛇和蝰蛇含有神经毒素。神经毒素的检测一般以大鼠膈膈神经和鸡颈二腹肌神经标本为依据。根据生理活动的不同,神经毒素可分为两大类,一类是突触前神经毒素,能抑制神经传导中乙酰胆碱的释放;另一种是突触后神经毒素,它可以与运动终板乙酰胆碱受体结合导致神经传导受阻。
1.突触后神经毒素
突触后神经毒素与肌肉运动终板上的烟酰胺乙酰胆碱受体结合,阻止神经肌肉传递。其作用特点与箭毒相似,故常称为拟箭毒样神经毒素或箭毒型神经毒素。自从这种毒素首次被分离出来后的十年左右时间里,它已被许多人广泛研究,并且是研究最多的毒素之一。目前已分离出数十种此类毒素,并对其一级结构、空间结构和化学修饰性质进行了大量研究。突触后神经毒素广泛存在于眼镜蛇和海蛇的毒液中,而这两种蛇毒几乎都含有一种以上的突触后神经毒素。蝰蛇科和响尾蛇的毒液在极少数物种中含有突触后神经毒素。
共同的物理和化学性质
突触后神经毒素不仅在结构上非常相似,而且在功能上也几乎相同,可归纳为以下几个方面:
1、结构相似,突触后神经毒素是一种小分子单链蛋白,不含糖和其他非蛋白成分,一般含有6070个氨基酸残基,45对二硫键。根据分子量和二硫键数目,突触后神经毒素可分为I型和I型神经毒素两类。 I型神经毒素又称短链神经毒素,由6062个氨基酸残基组成,含有4对链内二硫键; I型神经毒素又称长链神经毒素,由约70个氨基酸残基和5对链内二硫键组成。一般可用一个式子来表示某一分子的特征,如61-4,表示该毒素由61个氨基酸残基组成,含有4对二硫键。前田等。 (1974) 分离出一种特殊的突触后神经毒素,它有55 个氨基酸残基和5 对二硫键。毒素的部分氨基酸序列已经确定。第二和第三两个硫键之间的氨基酸序列中有一个Ala-Ala-Thr序列,与长链神经毒素对应的位置相同。根据二硫键的数量和氨基酸序列,该毒素属于长链神经毒素。无论是两类突触后神经毒素中各毒素之间,还是两类神经毒素之间,大部分氨基酸序列是相同的,二硫键的配对在两类突触后神经毒素中也是严格不变的,这方面将在第15 章的第1 节中详细讨论。
具有相同理化性质的突触后神经毒素几乎都是等电点在9-10之间的碱性蛋白质,由于分子量小但二硫键多,这些毒素的理化性质非常稳定,对耐热和化学试剂。即使在沸水浴中加热也不影响它们的活性。它们没有任何酶活性。积极的。后一个特征也有例外。有人从半环蛇的毒液中分离出三种具有PLA2活性的突触后神经毒素。由于PLA2 活性较弱,这些神经毒素过去未被发现。用更高剂量的毒素和更灵敏的方法测定,它们确实具有PLA2 活性。哈维等人。 (1980)研究了它们的PLA2活性与突触后神经毒素毒性的关系,发现其中有一种叫做磷脂酶A的成分!其神经毒性与PLA2 活性有关。毒性降低;而另外两种成分磷脂酶A3和鳞脂酶A4,其PLA2活性与神经毒性关系不大。
相似的药理特性无论是长链神经毒素还是短链神经毒素,它们都与运动终板上的烟酰胺型乙酰胆碱受体结合,阻止神经肌肉去极化,导致神经传导阻滞。它们具有与箭毒相似的作用,因此突触后神经毒素也被称为伪箭毒样神经毒素。突触后神经毒素虽然与箭毒具有相似的作用,但它们之间仍有许多区别:突触后神经毒素只能作用于烟酰胺型乙酰胆碱受体,结合慢,解离慢;但箭毒不仅可以与上述受体结合,还可以与粘多糖和其他蛋白质结合,如乙酰胆碱酯酶。与箭毒相比,突触后神经毒素与乙酰胆碱受体的结合更强,产生的生理效应也更强。例如,分子量为682的D-箭毒安眠药氯化物对小鼠的半数致死量为200ug/kg,分子量为70008000的突触后神经毒素的半数致死量为50ug/kg对于老鼠。测定时全部采用静脉注射进行,以摩尔计,突触后神经毒素的致死活性是箭毒的1540倍。静脉内和腹腔内注射突触后神经毒素同样有效,皮下注射效果较差。就Cobrotoxin 而言,皮下注射的毒性比其他注射剂低30%。对于箭毒,皮下注射的效果仅为其他注射的六分之一。
突触后神经毒素对受体的影响非常强,因为它们的解离常数为10-1。 ~ 10~11之间。测量是在模拟生理条件下进行的,所用材料为电鱼放电器官的细胞碎片,细胞碎片与受体结合,体系中无乳化剂。强结合的本质是亲水力和氢键。通过化学修饰可知,毒素与受体的结合不通过二硫键的相互作用。神经毒素与可溶性受体的作用不受高浓度NaCl(1.5mol/L)的干扰,说明结合不是静电作用,两类毒素的解离常数相同。已知长链毒素与受体的亲和力与温度有关,低于11亲和力迅速下降。一方面,这是由于疏水相互作用的减少;另一方面,随着温度降低,毒素与受体的结合速度降低,而解离速度影响不大。结合过程所需的热能解释了毒素或受体或两者的构象变化。在低温下,毒素无法完成这种构象变化。温度依赖性可以作为受体分离的条件,受体可以在室温下作为配体受到毒素的作用。
行亲和吸附,低温可以解离。突触后神经毒素的功能主要是阻断乙酰胆碱受体,但Denis等(1980)发现a-Bun- garotoxin和oc-Cobrotoxin还具有加强肺、睥和肾中鸟苷酸环化酶活性的作用。但对肝、肌 肉和脑组织中该酶的活性没有影响。如果对这两种突触后神经毒素进行还原和烷化处理, 上述作用消失。氧化解毒处理也能解除它们对环化酶的影响,但如用二硫赤藓糖处理只能 降低它们的作用。突触后神经毒素对环化酶的作用可能与它们改变了鸟苷酸环化酶的氧 化还原状态有关。因为改变该酶的氧化还原状态可以调节酶的活性。另外,Miller(1977) 发现某些突触后神经毒素能抑制由于病毒(Semliki Forest)感染大田鼠幼鼠肾纤维细胞而引起的鼠疫的产生。
(二)两类神经毒素之间的差别
虽然长链神经毒素和短链神经毒素在一级结构上相似,但在水溶液中,在没有配体的 情况下,二者的三维结构是不同的。不但如此,两类神经毒素还存在着下列差别:
1.免疫方面每类神经毒素的抗血清只能中和本类的神经毒素,在本类范围内无选 择性,但不能与另外一类神经毒素作用。也就是说,同类神经毒素抗原性相同,不同类神经 毒素的抗原性不同。神经毒素、心脏毒素和细胞毒素的抗原性都不同。每类毒素至少有3 个抗原决定簇,但各自都不同。
2.圆二色性两类神经毒素在圆二色性方面差异很大。如图3-12-1所示,它们的 C. D光谱,与标准蛋白质光谱相差很大,它们的a_螺旋、b-折叠和自由松散结构的含量都 不易测定。
3.稳定性尽管两类神经毒素对热都稳定,只能 被尿素和盐酸胍做可逆性的变性,但对其他处理如冷 冻和化学修饰产生的反应是不同的。在冷冻干燥处理 方面,长链神经毒素经冷冻干燥,本来单体的毒素可以 因冷冻干燥而产生微量三聚体、5%的二聚体,其余为 单体。如果是短链毒素,经冷冻干燥可以产生大量的二 聚体、三聚体和多聚体。在化学修饰方面,化学修饰对 短链神经毒素的活性影响很大,但对长链神经毒素的 活性影响较小,还没有发现修饰影响长链毒素的活性 而不影响短链毒素活性这种现象。也就是说长链毒素 比短链毒素对化学修饰剂反应要迟钝些,凡是能修饰 长链毒素的修饰剂都能够修饰短链毒素,反之就不一 定了。
4.对受体的结合两类毒素与受体结合的解离 常数在 pH7. 4、2(TC 时都为 5. 7X10-10mol/L〜8. 2X 10-9mol/L,但复合物结合的动力学不同。短链毒素与 受体结合的速度比长链毒素快6倍〜7倍,解离速度 也快5倍〜9倍。短链毒素阻滞神经-肌肉传导可以用 新斯的明洗涤而得到可逆的缓解,但长链神经毒素却 没有这种性质,这可能因为前者与受体结合后解离快 引起的。一般地说长链神经毒素的作用是不可逆的,不能被抗血清中和,而短链神经毒素 是可以被抗血清中和的。由于解离常数相同,所以二类神经毒素可能作用于受体的同一区 域。为了与该区域结合,神经毒素构象必须发生改变,根据动力学研究,长链神经毒素构象 改变所需的时间较长。结合到受体上的神经毒素的结构与自由态的结构完全不同,自由态 毒素分子内部的氨基酸在结合后与受体相接触,即内部的氨基酸基团转到分子表面。
不但不同类型的神经毒素存在上述差别,相同类型的毒素之间也存在着一定的差别。 64-4型(含64个氨基酸,4对二硫键)的短链神经毒素较易结晶;60-4型的毒素也能产生结晶,但无论做多少实验都不能使61-4型的毒素产生结晶。所有的长链毒素都没有得到 结晶。
(三)分离和纯化
突触后神经毒素的纯化程序与所用的蛇毒有关。对于眼镜蛇毒,常用CM-纤维素、 CM-Sephadex或SE-Sephadex来分离;或将蛇毒经两次硫酸铵沉淀后,经Sephadex G-75 脱盐,冷冻干燥,再用CM-纤维素分离,梯度洗脱。有关突触后神经毒素分离纯化的报道 十分多,各种神经毒素分离的过程大致相似,没有必要一一介绍,读者可以查阅有关这方 面的综述文章。在这里主要介绍一下从浙江蝮蛇Pallas)蛇毒中分离突 触后神经毒素的过程,并对我国研究突触后神经毒素的现状做一些讨论。
突触后神经毒素一般存在于眼镜蛇毒和海蛇毒中,响尾蛇毒和蝰蛇毒极少含有这类 毒素,中国浙江产竣蛇Zia/w Pallas)蛇毒属于特例。Jiang等(1987)顺序地用 CM-Sephadex C50 层析、Superose 6HPLC 层析和 DEAE-Sepharose C16B 层析从该毒中 分离出一种突触后神经毒素,称为a-Agkistrodotoxinh-Agt)。用SDS-PAGE测定该毒素 的分子量为8 000士80,能够和抗a-银环蛇毒素的家兔血清产生沉淀反应。该毒素还能竞 争抑制银环蛇毒素对培养的肌细胞烟酰胺型乙酰胆碱受体的结合,IC9() = 2Xicr9 M。它还能抑制由 Carbachol 诱导的 Ca2+通过 nAchR 的内流,IC50 = 6X 10_8mol/L。Jiang 还对它结合受体的结合与解离速度进行了计算,由这两种常数计算出Kdzy.sxicr1。 mol/L0
我国学者对神经毒素的研究起步较晚,蔡景霞等(1980)用羧甲基葡聚糖C-25和氯化 钠直线梯度方法,对我国广西产眼镜王蛇蛇毒进行了柱层析分离,获得17个蛋白峰,测定 了 I7个蛋白峰的毒性和生理活性,结果有7个毒性峰,其中6个峰具有阻断神经-肌肉接 头传导的作用,另一个峰毒性很小,在相同剂量下也不表现上述作用。在6个峰中,组分 7、8、9、和11蛋白回收率比较高,具有显著的神经-肌肉传导阻滞作用。由于它们还含有磷 酸单酯酶、磷酸二酯酶、5'-核苷酸酶和核糖核酸酶等活性,孙欣等(1981)进一步用羧甲基 纤维素CM 32和葡聚糖凝胶G-50纯化了组分7、8、9和11。通过聚丙烯酰胺凝胶电泳鉴 定,它们均为单一蛋白且具有神经-肌肉阻遏作用,没有磷酸单酯酶等酶活性。随后测定了 它们的氨基酸组成和N-末端氨基酸。离体大白鼠膈神经膈肌标本和去神经大白鼠膈肌标 本实验和氨基酸组成测定结果表明,眼镜王蛇毒似乎既含长链神经毒也含短链神经毒,它 们的作用部位是在神经肌肉突触后膜,对突触前膜没有影响,都是突触后神经毒素。林南 琴等(I984)对4种突触后神经毒素之一的CM-9进行了结构分析,应用固相DABITC- PITC双偶合Edman手工降解法能对完整的还原S-竣甲胺甲基化CM-9从N端降解至 第51位。联结经双向纸层析电泳纯化的、用胰蛋白酶裂解肽段的顺序,完成含73个氨基 酸残基的全部顺序。CM-9的顺序与Joubert报道的泰国眼镜王蛇神经毒素的顺序基本相 似。整个顺序中有18%氨基酸残基不同,特别在C端有4个残基有很大差别,但与银环蛇 «-毒素C端的疏水性相似。
















